RENÉ VILLLANUEVA MALDONADO, México
¿Es posible definir al árbol?
RENÉ VILLLANUEVA MALDONADO, México
¿Es posible definir al árbol?
A veces las preguntas más sencillas, tienen las respuestas más complejas, y es que la definición que todos creemos tener del árbol, en realidad no corresponde para nada a la realidad, al menos en su verdadera dimensión. Resulta que hemos estereotipado la silueta y el tamaño del árbol, de acuerdo a nuestra asociación empírica con algunas pocas especies que integran nuestra cotidianidad, lo cual nos genera una noción ambigua y simplista acerca de la complejidad evolutiva que representa la expresión arbórea dentro del mundo vegetal. Tenemos infinidad de formas de árboles, desde Sequoyas de 110 metros de altura, árboles coloniales que se clonan hasta formar bosques enteros, algunos que empiezan su vida siendo enredaderas para luego convertirse en gigantes arbóreos, o árboles subterráneos que esconden sus ramas bajo el suelo para huir de los incendios estacionales, y que apenas asoman sus discretas hojas una vez que el fuego ha pasado.
Cuatro problemas escenciales a la hora de definir la identidad del árbol
El primero
es que nuestra definición clásica del árbol, es aquella que ha permeado
en la percepción general de todo el mundo, en donde el árbol
simplemente se describe como una planta leñosa que ramifica a cierta
altura, que mide al menos cinco metros y que posee un solo tronco... El
gran problema con esta definición es que su origen radica en los países
de clima templado, cuya diversidad forestal, se limita a bosques
dominados por un puñado de especies arbóreas jamás podrá representar a
las aproximadamente 70.000 especies de árboles (Hallé, 2013) que representan la biodiversidad arbórea tropical.

Como
vemos dentro de esta definición estereotipada, la descripción del
árbol corresponde a una serie de formas que son clásicas de los países
europeos, Estados Unidos, o Canadá, donde debido a su latitud y longitud
geográfica se ubican dentro de climas templados. Cabe recordar que la
biodiversidad florística, particularmente de la composición forestal, es
extremadamente reducida en comparación con la de latitudes
ecuatoriales donde el clima es tropical. Esta explosión de vida en el
trópico, lógicamente se debe a la ausencia de restricciones
meteorológicas estacionales, al menos de forma prominente, y las
condiciones de temperatura y humedad son elevadas y constantes, a
diferencia de los países templados que tienen estaciones frías muchas
veces con nieve y heladas, secas, calurosas o lluviosas. Por ejemplo en
1968 POORE, contabilizó un total de 374 especies de árboles en una
pequeña zona de selva de 23 hectáreas al este de Malasia, dentro de la
reserva forestal Jengka. En contraste HORN en 1975 contabilizó un total
de 13 especies de árboles en todo el estado de Nueva Jersey, Estados
Unidos. La brecha que divide la exponencial biodiversidad de los
trópicos con los climas templados, parece casi infinita.
Nuestro segundo
problema a la hora de querer interpretar el árbol, es que tratamos de
justificar la delimitación de los caracteres que cada uno de nosotros
interpreta de un árbol, de manera distinta. Y es que para empezar la
palabra árbol no describe un término válido ante las ciencias botánicas,
sino que simplemente es una palabra coloquial que ha permeado
profundamente en la consciencia colectiva de la sociedad, sin que esta
cuestione o defina los límites de una palabra tan ambigua y vacía. Es
importante recordar que si bien nuestro lenguaje nos permite cuantificar
y clasificar nuestras necesidades comunicacionales, por definir
aquellas cosas que nos interesan tanto de nuestro entorno, como de
nuestra mente, en realidad el lenguaje constituye no otra cosa sino una
aproximación descriptiva de nuestras interpretaciones epistemológicas,
por lo que culturalmente es necesario que reconozcamos sus limitaciones.
Aspectos como el tiempo, la longitud, la masa o el peso, son unidades
de medida cuya construcción depende directamente de nuestras necesidades
culturales, sin embargo las medimos a partir de puntos arbitrarios que
convienen a nuestra comprensión práctica y funcional. Por otro lado,
tales conceptos desde un punto de vista meta-humano, simplemente no
existen ni aplican a la forma en que funciona la naturaleza. Recordemos
que el antropocentrismo que nuestra cultura enfunda en nuestras
interpretaciones, nos dan una falsa noción de realidad o de control
acerca de lo que nos rodea, sin embargo el mundo y la materia no
funcionan en nuestros términos culturales. La ciencia y las "verdades"
son temporales a cada siglo, década y momento histórico que definen los
cambios que sufre nuestra cultura, por lo que su validez constituye una
interpretación temporal casi siempre, no es estática, se descarta, muta,
se complementa, etc. El problema es que nuestro lenguaje, siempre
impregnado de nuestra temporalidad cultural, resulta ambiguo e
insuficiente para describir con amplitud de perspectivas, todos los
elementos que definen el mundo natural, entre ellos los árboles.
En
este artículo, no pretendo revelar ninguna verdad acerca de lo que es
un árbol, sino revelar algunos de nuestros vacíos culturales y tratar de
construir una interpretación más aproximada a una definición sobre la
entidad conceptual que en nuestro lenguaje llamamos "árbol".
El tercer problema
radica en que aquello que llamamos "tronco" y que define el órgano de
transición entre la copa y la raíz, que provoca la altura del árbol, en
realidad constituye uno de los más grandes errores de interpretación de
nuestro cliché arbóreo. Y es que resulta que el tronco de un árbol se
puede construir de distintas maneras que no siempre tienen que ver con
aquello que coloquialmente reconocemos.
 |
| Imagen: Tropical an Forests de F.Hallé, R. Oldeman... |
Para empezar debemos tener presente la próxima vez que queramos definir a un árbol a la Arquitectura Arbórea y
es que en realidad existe toda una disciplina botánica que se dedica a
estudiar las reglas por medio de las cuales las plantas se construyen a
través de diversas estrategias de organización de los caracteres
morfológicos, los cuales determinan la repetición de estrategias
construccionales que son mediadas tanto por factores endógenos
(genética, plasticidad fenotípica) así como los exógenos (restricciones
del medio ambiente, interacciones ecofisiológicas, sincronismo de etapas
de desarrollo u ontogenia). Se reconocen hasta el día de hoy 24
estrategias construccionales, llamadas modelos arquitecturales, de los
cuales les hablaré en posteriores artículos.
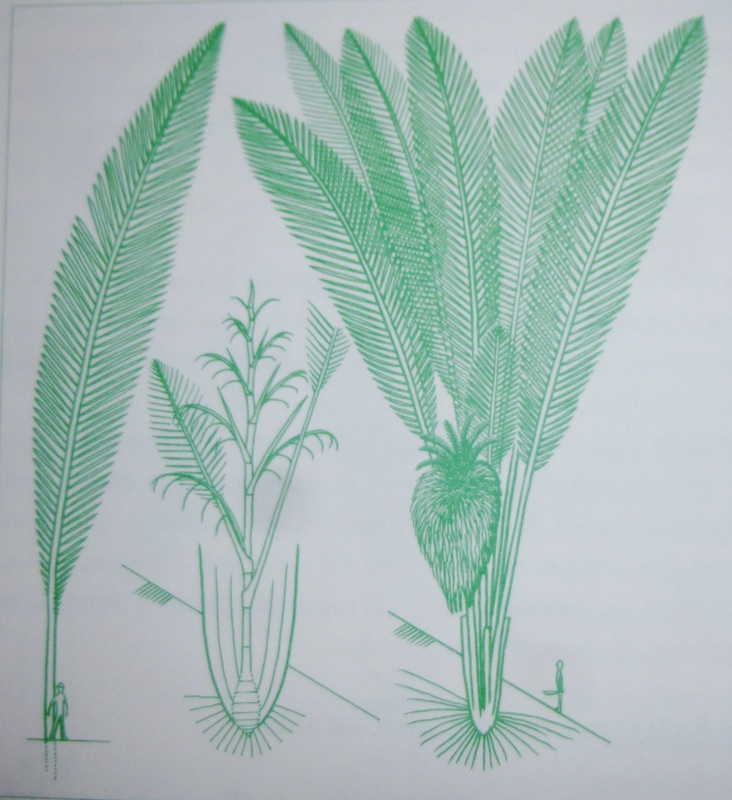
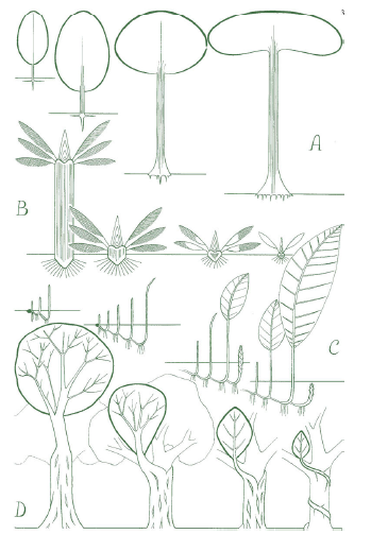
Por ejemplo; en el diagrama de la derecha, observamos distintas estrategias para construir un tronco.
La imagen A,
constituye la más común de las estrategias reconocibles para la mayoría
de las personas. Un tronco se construye bajo esta estrategia, ya sea
por medio de la creación de un tronco central que se construye por medio
de un líder apical (ejemplo Araucaria, Abeto, etc Modelo de Massart),
por relevo de ramas súper puestas que intercalan funciones y se
diferencian en un tronco a medida que maduran jerárquicamente dentro de
sus etapas de desarrollo (Ejemplo Acacia, olmo. Modelo de Troll, Modelo
de Champagnat, Modelo de Mangenot). Aunque me quedo corto de ejemplos,
esta es la estrategia de construcción del eje principal o tronco, más
reconocible en los árboles dicotiledóneos.
La imagen B,
nos describe una forma más básica de construir un tronco que se
construyen por la acción de la actividad de una única yema terminal o
meristemo, que por medio de la superposición de frondas u hojas,
desarrolla un tronco que no ramifica y cuyo crecimiento se da en un solo
sentido, generalmente vertical (ejemplo: palmera, Furcraea, algunas
especies de Cacao, etc, Modelo de Holtum, Modelo de Corner, Modelo de
Chamberlain)
La imagen C, nos describe algunos
árboles que de hecho carecen de un único tronco y cuya ramificación es
basal o a través de un leptomorfo como en el bambú, donde un eje
morfológicamente idéntico al aéreo, explora bajo la tierra su entorno y
del cual brotan ejes aéreos verticales u ortótropos. La identidad de
estos árboles se basa en el relevo de ejes que son equivalentes en
funciones y que conforman como si fueran grupos de un solo árbol.
Ejemplos son el Bambú, el gengibre, algunos tipos de hoja santa, género
Piper, palmeras como la datilera, etc, Modelo de Tomlinson y Modelo de
Bell.
La imagen D, nos describe una estrategia
asombrosa, que es la construcción de un tronco, por medio del
entrecruzamiento en forma reticulada y posterior fusión o anastomosis
del sistema de raíces, sobre un soporte ya sea viviente o no. Ejemplos
de esto son las higueras extranguladoras, o Ficus, Araliaceas o
Clusiaceas, Modelo de Rauh, Modelo de Attims.

El cuarto problema
que tenemos es la escala o el tamaño que debe de tener una planta para
ser considerada un árbol, ya que este paradigma, nos llevará a muchas
encrucijadas ecológicas, evolutivas, culturales, etc. Por ejemplo, la
escala de medición de los árboles, siempre parte de la medición de
nuestra escala humana. Empezamos a medir al árbol, a partir de donde
paramos nuestros pies, sin embargo debemos recordar que el árbol no
empieza donde están nuestros pies, sino que solamente medimos la parte
aérea del árbol. En ocasiones la parte subterránea representa una
longitud vertical mayor hacia el fondo de la tierra, que su parte aérea.
De esta forma, tenemos algunas especies desérticas con raíces
pivotantes freatófitas (que buscan agua de los mantos freáticos) cuya
copa o parte aérea puede medir dos o tres metros de altura, pero sus
raíces pueden alcanzar hasta 80 m de profundidad.
Otro problema
asociado al tamaño, es que algunos árboles, sólo adoptan el porte
clásico arbóreo, si las condiciones ecológicas permiten la expresión
morfológica de este. Por ejemplo, algunos encinos pueden ser gigantes de
más de 20 m, creciendo en los bordes húmedos de las laderas montañosas,
mientras que estas mismas especies son arbustivas en regiones más
secas. En casos aún más confusos, el cambio de condiciones ecológicas
puede determinar si una planta se convierte en un árbol, en una
enredadera, o en un arbusto.
Desde el punto de vista evolutivo,
la manifestación del porte arbóreo tampoco es exclusivo de una familia, y
una vez más, al estar ligado a reglas de expresión arquitectural; estas
las podremos encontrar tanto en plantas monocotiledóneas, como en
dicotiledóneas. Por lo tanto, la arborescencia, no depende de los grupos
filogenéticos a los que pertenecen las familias que la expresan, sino
que principalmente el porte arbóreo está condicionado con las
necesidades adaptativas que las especies requieren para sobrevivir en
determinados nichos ecológicos. La edificación requerida para que una
planta se convierta en un árbol, depende por si misma de la capacidad
que esta tenga en su potencial genético, y a su vez poder contar con los
recursos ecológicos necesarios para activar los mecanismos de
crecimiento, a través de satisfacer las necesidades ecológicas de la
especie.
Es por eso que en determinados lugares, algunos árboles
que crecen en sitios ricos en recursos, expresan su potencial arbóreo al
máximo de los límites capaces de presentar el potencial genético y
fisiológico de la especie. Por otro lado, esta misma especie creciendo
en un sitio con recursos limitados, probablemente altere algunas de sus
características morfológicas para ajustarse al medio que la sustenta.
Por ejemplo la expresión de caudex basales en algunas especies.

Imagen comparativa de una misma especie de árbol, Pseudobombax ellipticum, Modelo de Koriba, creciendo en condiciones ecológicas distintas, y expresando un desarrollo vegetativo diferenciado.
En la imagen de la izquierda se aprecia el desarrollo de un caudex
basal bien definido en el árbol, a partir de esa zona, el árbol almacena
recursos metabólicos. Al estar creciendo entre rocas, y el espacio y
recursos ser tan limitado, el árbol inhibe el desarrollo de troncos o
ejes principales, su ramificación es basal y la principal zona de
crecimiento es a partir del caudex. Los árboles desarrollan estructuras
acordes a las restricciones del medio físico como puede ser la gravedad,
por lo que si su posición es en una pendiente o acantilado fuertemente
pronunciado, el árbol puede orientar su crecimiento en favor de las
fuerzas de gravedad, es decir, expresar un hábito de crecimiento
péndulo, o semi péndulo, con la orientación de las hojas re dirigidas
hacia la fuente luminosa.
En la imagen de la derecha, vemos a la
misma especie de árbol creciendo en un suelo poroso, con espacio
suficiente, humedad y buen drenaje. Esto activa los mecanismos de la
expresión arquitectural arborescente, por lo que el árbol desarrolla una
forma más a fin a nuestro estereotipo arbóreo.
En este ejemplo se muestran tres imágenes un género de plantas sudamericanas llamado Vellozia.
En la imagen de la izquierda arriba, observamos como estas plantas,
emparentadas tanto con los lirios que decoran nuestros floreros y
jardines, cómo con los Pandanos asiáticos y africanos, han desarrollado
estrategias ecológicas arborescentes, a partir de un ancestro más
ordinario como una planta con bulbo subterráneo, como el que se aprecia
en la imagen central inferior. Las estrategias evolutivas relacionadas a
la arborescencia, una vez más nos demuestran ser estrategias
adaptativas condicionadas a las circunstancias ambientales.
A que extremo es necesario ejemplificar las limitaciones lingüísticas y culturales de la indefinición del árbol?
Bien, aquí tenemos dos fotografías de un mismo árbol procedente de África, llamado Parinari capensis.
El asunto, es que ambos son la misma especie pero pertenecen a
diferentes subespecies, siendo así que la subespecie de la foto de la
izquierda, Parinari capensis incohata, es un pequeño árbol que
vive en pastizales y que da sus flores y frutos en lo alto de sus ramas.
Mientras tanto, la foto de la derecha: Parinari capensis capensis, es un árbol subterráneo!
Así
como lo dije, un árbol morfológicamente idéntico a su pariente
incohata, pero en donde sus ramas se encuentran bajo la tierra y este
apenas asoma sus hojas, flores y frutos por encima del nivel del suelo.
Esta
adaptación de trasladar la estructura arbórea, abajo de la tierra,
resulta en una condicionante ambiental muy importante. El sitio donde
esta subespecie vive, constantemente es arrasado por incendios, por lo
que gastar energía en crear madera y estructuras aéreas, sería un
desperdicio, debido a que estas se quemarían constantemente. Mejor el
árbol se ha enterrado, y si el incendio pasa; este sólo habrá quemado
pequeñas hojas y tallos, en lugar de toda la estructura leñosa. Esta
subespecie subterránea es extremadamente longeva, y se sabe de
individuos que tienen hasta 13,000 años de antigüedad, y que conforman
increíbles bosques subterráneos.
Bosque subterraneo de Parinari capensis capensis
La
ambigüedad del término árbol, trae errores muy comunes, incluso por
parte de personas expertas en árboles. Uno de ellos es el desligar a las
plantas como palmeras, yucas, dracaenas, entre otros, con los árboles,
al calificarlas como hierbas gigantes. Esto, evidentemente constituye
una equivocación mayúscula, ya que las monocotiledóneas también entran
dentro del reino arbóreo. Se dice que una de las diferencias, es que
estas plantas no compartimentalizan como lo hacen los árboles
verdaderos, sin embargo esto no es así. Si bien, la mayoría de las
monocotiledóneas no poseen la misma configuración anatómica que los
árboles convencionales, las monocotiledóneas sí compartimentan, aunque,
debido a la ausencia de cambium (la mayoría), sólo se activan las 3
primeras barreras del CODIT, y no la última, aunque en algunas como en
el género Yucca, la compartimentación completa, con la última barrera,
si llega a ocurrir, esto no debido a la acción de un cambium como tal,
sino a la desdiferenciación de tejidos parenquimáticos con potencial
meristemático. Algo que también ocurre en las dicotiledóneas. Las
monocotiledóneas y las dicotiledóneas no son tan distintas como se
piensa, en realidad son muy equivalentes.
La identidad arbórea de
las palmas, Dracaenas, Yuccas, etc, radica más en su relación
arquitectural con respecto a su configuración espacial dentro de nichos
ecológicos ecosistémicos, que en el hecho que de que su forma nos
resulte o no, familiares. En el caso de las palmeras, vemos 4 modelos
arquitecturales asociados a las características construccionales de sus
especies.
La arquitectura arbórea y su rol dentro de la determinación construccional arbórea
Las especies monocárpicas o hapaxánticas (que florean una sola vez y después mueren) corresponden al modelo de Holtum.
|
Corypha utan.
El modelo de esta palma, denominado Holttum, en honor al profesor
Inglés, Richard Erick Holttum, se caracteriza por crecer en un único
sentido, por medio de la actividad de un solo meristemo primario, tras
lo cual, este se modifica y se transforma en una inflorescencia masiva
que agota los recursos metabólicos de la planta hasta matarla, además de
que la planta no posee vías alternas para ramificarse, por lo cual su
muerte es inevitable.
Este modelo es común en monocotiledóneas, sin embargo también existe en dicotiledóneas.
|
|
Perteneciente al modelo de Holttum, encontramos a la planta con las hojas más grandes del planeta: la palmera africana Raphia regalis,
cuyas hojas alcanzan los 25 metros de longitud. Ilustración tomada del
libro: Plaidoyer pour l'arbre, cuyo autor es Francis Hallé.

Aquí vemos un árbol dicotiledóneo perteneciente al modelo de Holttum: Spathelia sorbifolia,
originaria de Jamaica. Se trata de un árbol monocárpico que alcanza
hasta 20 metros de altura, antes de que se imponga la inflorescencia y
acabe con la vida del árbol.
El
segundo modelo que vamos a ver asociado a las palmeras, pero que está
presente también a los árboles dicotiledóneos es el modelo de Corner. El
nombre de cada modelo, es el apellido de algún botánico que haya
contribuido de manera significativa al entendimiento de la arquitectura
vegetal.
|
Arriba: Bikkia macrophylla
Abajo: Ceroxylon quindiuense, la palma más alta del mundo, originaria de Colombia. Alcanza los 80 metros de altura,
El
siguiente modelo que vamos a analizar, igualmente muy afín a las
monocotiledóneas arbóreas, es uno que en este caso si está ramificado.
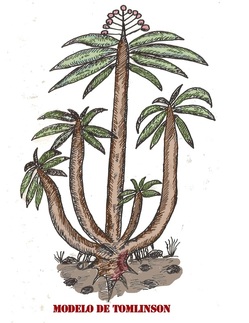
Se trata del modelo de Tomlinson, el cual se
caracteriza por tener múltiples ejes, cuya construcción individual,
podría ser idéntica a la del modelo de Holttum o Corner. Sin embargo,
este modelo, ramifica de manera basal.
Modelo de Tomlinson
Phoenix dactylifera
|
Oenocarpus distichus. En
esta palma, se representa el modelo de Corner, cuya única diferencia es
que es policárpico (que florea múltiples veces) Su floración es
lateral, y no terminal como en Holttum, por lo que la planta no detiene
su crecimiento por la floración. No ramifica este modelo. Aunque es
común, en monocotiledóneas, también lo es en dicotiledóneas como el
árbol de papaya.
Árbol de papaya Carica papaya
|
A
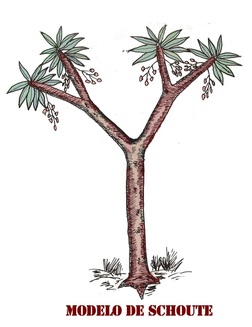
continuación veremos un modelo muy particular dentro del mundo vegetal,
puesto que define a las plantas que ramifica por división de meristemo,
produciendo el fenómeno de dicotomía verdadera. El resto de las
plantas, ramifican por medio de meristemos (yemas) laterales
secundarios. Se trata del modelo de Schoute, y para ejemplificarlo, usaremos a una palma icónica de este modelo: Hyphaene thebaica.
Exactamente se trata de una palmera ramificada a modo de árbol
dicotiledóneo, y que en este punto comenzará a confundirnos aún más con
el término "árbol".
Hyphaene thebaica
Modelo de Schoute
Después de ver los modelos de las palmeras, y comprobar que su
identidad arbórea es real, veremos otro modelo comparativo entre las
monocotiledóneas y las dicotiledóneas del mundo arbóreo:
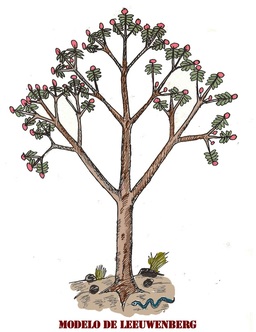
El modelo de Leeuwenberg
Este
modelo arquitectural, comienza comportándose en un inicio como el
modelo de Holttum. Sin embargo, tras la floración, en lugar de que muera
la planta, este modelo produce 2 o más ejes contiguos a la zona de
floración, los cuales son simpodios o módulos sobrepuestos sobre el
anterior que floreció. Su morfología es igual en todas sus partes ya que
se comporta como si fuera una repetición o reiteración de la unidad
básica del modelo. La copa de este modelo, suele ser muy geométrica y
predecible en su secuencia de ramificaciones. El modelo de Leeuwenberg
es tan común en monocotiledóneas, como en dicotiledóneas, excepto en
palmeras.
Senecio praecox, de México. Dracaena cinnabari, de la isla de Socotra en Yemen.
Una dicotiledónea. Una monocotiledónea.
Los árboles en el registro fósil
Equisetum arvense, una planta de hasta 50 cm de altura. Su origen evolutivo es muy antiguo.
|
Paisaje boscoso del periodo Carbonífero hace 359 millones de años
Calamites:
un árbol propio del periodo carbonífero que alcanzaba hasta 30 metros
de altura, sin embargo era muy similar a lo que hoy son el género Equisetum, o colas de caballo. Modelo de Bell.
|
|
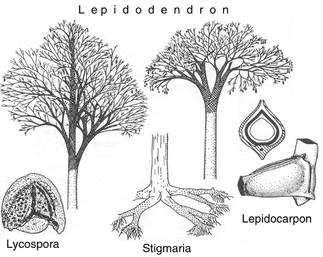
Lepidodendron: Una especie extinta del carbonífero que llegó a medir hasta 35 metros. Muy emparentada con los actuales Lycopodios.
|
|
|
Aquí se observan las hojas o microfilos del Lepidodendron, así como sus estróbilos.
|
|
Abajo
de Lepidodendron, vemos dos fotos comparativas de los actuales
Lycopodios y Lycofitas en general. Comparten características evolutivas
muy similares, pero los lycopodios son actualmente pequeñas plantas
epífitas o rastreras de los bosques húmedos.
|
Tratemos
de reconocer a los árboles de acuerdo a su origen morfológico, según el
registro fósil. Si bien a primera vista la mayoría de nosotros no
dudaría de la identidad arbórea de muchos árboles del periodo
carbonífero, en realidad pertenecieron a familias que hoy en día nos
parecerían completamente alejadas a lo que las definiciones
convencionales de árbol tenderían a encasillar, sin embargo fueron
árboles de hasta 60 m. Ya fuera como Helechos o equisetos, plantas que
hoy cultivamos en maceta y jamás reconoceríamos como árboles, en el
periodo carbonífero lo fueron.
De ahí que debamos trasladar nuestra
definición de árbol, más a una estrategia, evolutiva, ecológica, o
fisiológica, que a una forma definida.
Equisetum arvense: Una planta muy relacionada al extinto Calamites, de porte herbáceo que mide en promedio 50 cm de altura. Modelo de Bell.
|
Conclusiones
Como
hemos visto, es sumamente ambiguo definir al árbol desde una
interpretación cultural lingüística, ya que parte de un estereotipo, el
cual queda anulado cuando se le da la dimensión global de la diversidad
florística arbórea del planeta, por lo que ¿cómo podríamos aproximarnos a
la definición más acercada a un árbol?
En primer lugar debemos
empezar a reconocer que partimos de plantas que ocupan los nichos
ecológicos dominantes, dentro del sistema de clasificación de estratos
forestales. Sin embargo esta propiedad basada en una característica
adaptativa, no posee barreras filogenéticas bien delimitadas: El árbol
es como definir el arte, cualquier definición, en general suele ser
inexacta e insuficiente. Sin embargo el arte se define así mismo a
través de sus artistas y sus obras, más que por un estereotipo temporal
que rápidamente caducará ante la transmutación cultural que
constantemente sufre a través de nuevas tendencias, movimientos,
tecnología, etc. Así mismo hay que ver el árbol, quizás no tanto como
algo definible en uno o dos renglones. El árbol se define por su
diversidad de especies y de formas, por su relativo amorfismo, por
nuestra limitada interpretación del mundo, por nuestra relativa
realidad, por nuestro lenguaje insuficiente.
Reconocer o no una
definición, depende de cada interpretación, de este modo se comprueba
que el árbol no es un ser definible. Sin embargo podemos quedarnos con
el tamaño, la forma o el soporte a sí mismo, si es que no queremos
rompernos la cabeza con confusiones... eso hasta que descubramos que
muchos de ellos comienzan las primeras etapas de su vida como
enredaderas. O por su tiempo: aunque luego tengamos que reconocer que
muchos árboles comienzan su vida como arbustos y que tardan más de 100
años en comenzar a alcanzar su porte arbóreo, por lo que en nuestra
escala del tiempo, tan desfasada a la de los árboles, ese organismo
jamás será reconociblemente un árbol durante el periodo de vida de un
ser humano.
Ejemplos comparativos de árboles actuales
Foto Izquierda: Dracaena cinnabari, Socotra, Yemen.
Foto abajo: Dracaena draco, Tenerife, España
Modelo de Leeuwenberg
Corypha utan, Sudeste asiático y Australia.
Modelo de Holttum
Ceroxylon quindiuense, Palma de cera en Colombia, altura de hasta 80 m
Modelo de Corner
Aloe dichotoma: Namibia
Modelo de Leeuwenberg
Puya raimondi, Perú y Bolivia
Modelo de Holttum
Dendrocalamus giganteus: Bambú gigante, Tailandia e Indonesia
Modelo de Bell
Alpinia carolinensis: Gengibre gigante, Sudeste asiático
Modelo de Tomlinson
Dendrosenecio kilimanjaro, Monte kilimanjaro, Tanzania
Modelo de Leeuwenberg
Beaucarnea gracilis, Tehuacán, Puebla, México
Modelo de Leeuwenberg
|
Yucca brevifolia, México
Modelo de Schoute
|
|
Cycas revoluta, 400 años. Japón
Modelo de Corner?
Cycas tansachana, Tailandia
Modelo de Chamberlain
Terminalia catapa. India.
Modelo de Aubreville
Magnolia grandiflora, EUA.
Modelo de Fagerlind
Pereskia lychnidiflora, México
Modelo de Koriba
Carnegia gigantea, México-EUA
Modelo de Rauh
Euphorbia ingens, Malawi
Modelo de Rauh
Abies alba, España
Modelo de Massart
|
Aniba roseodora, Venezuela
Modelo de Rauh
|
|
Bibliografía Consultada
-Tropical Trees and Forests: Francis Hallé, P.B. Tomlinson, Roelof Oldeman
Plaidoyer pour l'arbre: Francis Hallé
The CODIT Principle, English Edition. Dirk Dujesiefken, Walter Liese.
A New Tree Biology: Facts, Photos, and Philosophies on Trees and Their Problems and Proper Care. Alex Shigo.
Plant form: Adrian D. Bell
ESSAI SUR L'ARCHITECTURE ET LA DYNAMIQUE DE CROISSANCE DES ARBRES TROPICAUX-
-Francis Hallé, Roelof Oldeman
Tree Architecture: P.B. Tomlinson
-----
 El Archipiélago Canario formando el frente contra el que chocan las nubes de los vientos aliseos cargados de humedad...
El Archipiélago Canario formando el frente contra el que chocan las nubes de los vientos aliseos cargados de humedad...